A digital head for Acanthostega

What has 16 fingers and a digital skull? Acanthostega, that’s what!
Acanthostega was one of the first limbed (rather than strictly “finned”) vertebrates, living around 365 million years ago in the shallow waters of modern day Greenland. Imagine something that looked roughly like a two foot long cross between a salmon and a salamander. As an odd evolutionary quirk, perhaps representing early experimentation in tetrapods (limbed vertebrates), Acanthostega had eight fingers on each of its hands. This critter is known from the nicely preserved fossils of several individuals, which has deservedly given it the title of “iconic” for our understanding of the transition from water onto land, for everything from feeding to locomotion.

{kind=link}
Of course, we paleontologists are never satisfied with beautiful fossils. There always seems to be something that isn’t quite preserved, or is preserved in the wrong orientation for easy viewing, or is crushed in such a way as to render opinions on anatomy equivocal. Frustratingly, these kinds of details can be important for nailing down the finer points of evolutionary relationships as well as behavior and function. During the last two decades, non-invasive medical imaging techniques have been adapted for digital preparation and reconstruction of many fossils. Thus, it is increasingly easy to pull out previously inaccessible details of ancient anatomy.
Laura Porro and colleagues just published a paper detailing their new digital reconstruction of the skull of Acanthostega, based on CT (computed tomography) scanning of three different specimens. Each fossil had some degree of crushing or incompleteness, but together they essentially added up to a whole skull. The trick, of course, was getting them to all fit together.
The first step is to digitally remove the rock from the fossils. This process involves making a judgment in the grayscale CT scans as to where one ends and the next begins. When there is high contrast (perceived by us humans as sharp color differences in the scan), this can be done fairly automatically by the computer; in less clear cases, a human has to make this decision. Anatomical knowledge as well as a “feel” for the scan are key.
Although we often think of a “skull” as a single unit of bone, the head is in fact made up of a number of separate but interlocking bones. Thus, each bone digitally was separated from its neighbor. The result is a bunch of colored bones floating in space, which are in their original position as preserved but not necessarily life position.
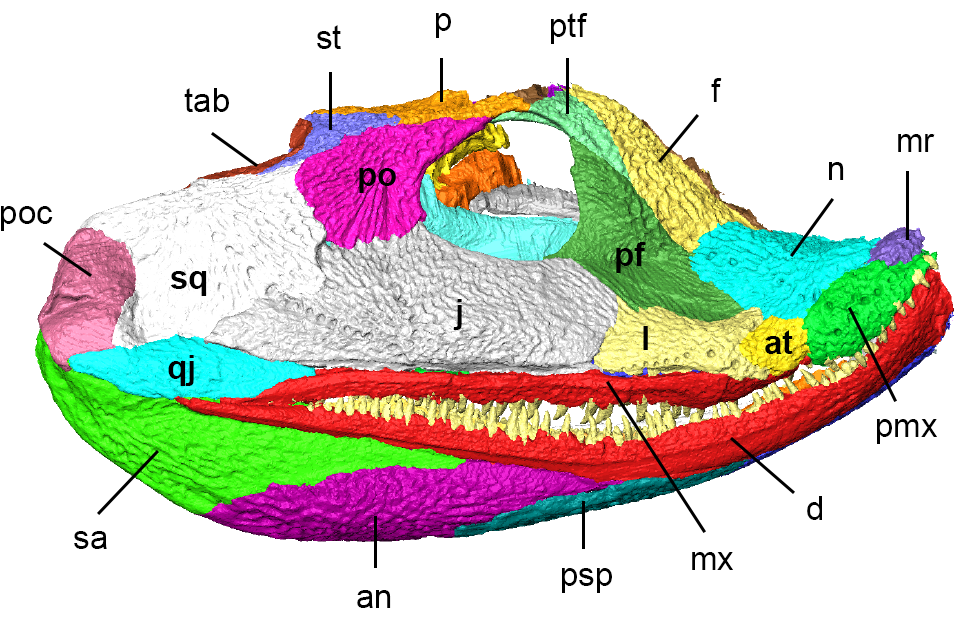
The next step is to fill in missing pieces and restore the skull to its life condition. Missing or incomplete bones could be replaced by scans from more complete fossils, scaled appropriately. Removing the post-death displacement of bones was a little more challenging. For animals with loosely-knit skull bones, individual bones tend to “sag” relative to each other after death, squishing some parts that weren’t originally squished. In the fossils, this can be evidenced through big gaps that obviously weren’t there in life, or through disturbingly overlapping bones that once again don’t look like anything in a living animal. Through digital manipulation, each bone can be scooted back to its life position relative to the other bones. Even in the best case this is a long and tedious process, and the authors clearly sank a lot of work into getting it right with Acanthostega. The result (below) is pretty dramatically different from the “as-is” fossil.

Although this isn’t the first attempt to “undeform” the skull of Acanthostega (previous pen-and-paper versions have been published), it is perhaps the best supported and most constrained by the fossil data. A quick glance at the original fossil as well as the digital reconstruction shows a number of difference between the two–the overall shape of the snout, position and size of the eye sockets, angulation of the back half of the skull relative to the front half, etc. The overall skull shape wasn’t the only thing that was newly revealed–the anatomy of some bones closely related to feeding (the quadrate bone, connecting the upper part of the skull to the lower jaws, for example) are also known for the first time.
There are no massive reinterpretations of the life, appearance or evolutionary history for Acanthostega in this paper, but that is okay. Nonetheless, descriptions of previously unseen bones and relationships between bones help generate refined hypotheses for future testing. For instance, Porro and colleagues further confirmed a previously suspected gap between bones on the top of the snout called the midline rostrals and nasals. This gap wasn’t immediately visible on the fossils themselves, but was unavoidably open when reassembling the skull on the computer. Some other early tetrapods also show this feature, and thus it might be handy for clarifying evolutionary relationships between these species.
Porro and colleagues also were able to describe the shape and texture of the contacts between individual bones (typically called sutures) at a new level of detail. The nature of these sutures–whether they are tightly linked from bone to bone, broad, narrow, sinuous, or whatever–can be correlated with feeding style. Something chowing down on actively wriggling prey might want to have a beefy skull with sturdy attachments between bones, for instance. If different parts of the skulls are used differently, this might also be reflected in the sutures. Some arrangements show resistance to being pushed together or pulled apart, and others show resistance to twisting. In the case of Acanthostega, it is hypothesized that the front of the skull was used for nabbing live prey (based on the teeth as well as the sturdy connections between bones on the front and roof of the skull). In contrast, the mid-section (as shown by the more loosely connected bones on the roof of the mouth and in the cheek) was more of a “stabilizer”, holding prey in place as Acanthstega swallowed. These interpretations can now be worked out in more detail using computer models. Overall, the evidence from this and other studies points towards Acanthostega feeding underwater (perhaps as a suction feeder), rather than on land.
The field of digital paleontology is no longer as new and shiny as it once was–indeed, CT scanning is now a fairly standard technique for many scientists. But, the case of Acanthostega once again shows how digital fossils have moved from “gee whiz we scanned a specimen” into “here’s a whole bunch of new anatomy and behavior.” Although it would have been pretty cool to see this animal devour live prey, I just as much like the fact that it’s now devouring internet bandwidth as I read the open access paper and soak in the beautiful imagery!

Soapbox Addendum
On a final note, I particularly commend the authors for including this phrase in their paper:
It should be noted that the 3D model represents our hypothesis of the reconstructed skull of Acanthostega, based on available specimens, scan resolution and personal interpretation.
When I read that sentence, I practically wept sweet tears of replicable science joy. As I have stated elsewhere, even in the best cases CT reconstructions require a certain level of finesse, practice, and personal judgment, with the strong possibility of alternative interpretations. It is wonderful to see that fact acknowledged in the literature–this is pretty rare otherwise. Additionally, although I strongly prefer that raw scans themselves be made available whenever possible, the authors have done a good thing by indicating that the scans are archived at the institution housing the original fossil specimens. I’m glad that there are more and more people thinking about reproducibility in digital reconstructions.
Citation
Porro LB, Rayfield EJ, Clack JA (2015) Descriptive anatomy and three-dimensional reconstruction of the skull of the early tetrapod Acanthostega gunnari Jarvik, 1952. PLOS ONE 10(3): e0118882. doi:10.1371/journal.pone.0118882